Projects
Activation and evolution of Vγ9Vδ2 T cells and the physiological significance of butyrophilins
Most T lymphocytes are MHC-restricted "conventional" T cells. These express a T-cell antigen receptor (TCR) containing a TCR α and a β-chain (αβTCR) that binds complexes of (foreign) peptides and "antigen-presenting" MHC class I and II gene encoded cell surface molecules. These T cells and the antibody-producing B cells are carriers of the adaptive immune response. The "non-conventional" T cells also express TCR, but show typical properties of cells of the natural immune system. They can be activated very quickly, and their TCR recognizes disease-associated molecular patterns rather than pathogen-specific antigens. In many cases, they are not (only) used to fight pathogens or cancer cells, but also to control the immune response or maintain the barrier function of tissues. We investigate the mechanisms and evolution of this antigen recognition and the activation of such cells.
Non-conventional T cells include αβ T cells, which are not MHC-restricted, as well as γδ T cells, which are defined by their TCR, which contains a TCRγ and a TCRδ chain. There are two main groups of non-conventional αβ T cells, MAIT cells and iNKT cells. Their TCR do not bind peptide-MHC complexes, but complexes of MHC class I-like molecules (MR1 or CD1d) and folic acid metabolites (MAIT cells) or (glyco)lipids, respectively. The γδ-TCR, on the other hand, binds a very wide range of structurally very different antigens and many unknown ligands. Like B cells and αβ-T cells, γδ T cells are found in almost all jawed vertebrates (Gnathostomata).
A special feature of many non-conventional T cells is that the use of certain TCR-V regions correlates with their cell function and localization. This differs from the TCR of MHC-restricted αβ-T cells, for which show no such correlation and are mainly found in lymphatic organs. An example of such T cells defined by the TCR genes are the Vγ9Vδ2 T cells we studied, whose TCR contain Vγ9 and Vδ2 gene-encoded V regions. Like many non-conventional αβ T cells, they recognize metabolites of microbial origin or metabolites that accumulate in stressed or tumor cells.
1–5% of peripheral blood T cells are Vγ9Vδ2 T cells. They express Vγ9Vδ2 TCR and recognize so-called phosphoantigens (PAg), which are phosphorylated metabolites of isoprenoid synthesis. Vγ9Vδ2 T cells attack tumor cells and body cells infected with PAg-producing pathogens and spread massively in infections such as malaria or some bacterial infections. The PAg do not bind directly to the Vγ9Vδ2 TCR but must be "presented" by cell surface molecules. However, this presentation does not occur by binding of the PAg to the extracellular domain of the MHC-like molecules, but by binding to the intracellular domain of the cell surface molecule BTN3A1. After binding to BTN3A1, an activating complex forms is formed. This contains BTN3A1, the structurally similar BTN2A1 molecule and non-PAg-binding BTN3 molecules (BTN3A2 and/or BTN3A3). This complex then acts as a T-cell-activating ligand of the Vγ9Vδ2 TCR. The Vγ9Vδ2 T cell thus recognizes changes in cell surface proteins that are induced by metabolites within the cell.
Phylogenetically Vγ9Vδ2 T cells appear together with placental mammals (Eutheria), but are no longer present in many species such as mice, so there is no small animal model of PAg-reactive cells. However, we were able to show that not only humans and other primates possess functional Vγ9Vδ2-TCR and BTN3A genes and that these are also found in alpacas (Vicugna pacos) link. The alpacas Vγ9Vδ2 T cells also react to PAg. Comparing BTNs and Vγ9Vδ2 TCR of both species allowed us to make predictions about certain properties that are essential for the function of these molecules. These hypotheses were experimentally tested by expressing modified BTN3 genes in BTN3-deficient cells and analyzing these cells for various features such as the ability to "present" phosphoantigens, the localization of BTN3 constructs in the cell, biochemical properties and formation of oligomers, and the interaction with the BTN2A1 molecule.
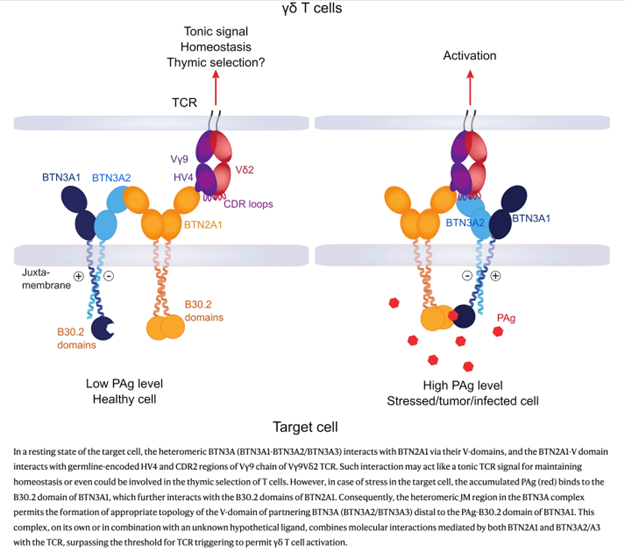
Figure from Phosphoantigen recognition by Vγ9Vδ2 T cells.
Herrmann T, Karunakaran MM. Eur J Immunol. 2024 Nov;54(11):e24510
These experiments led to a new model of PAg-induced Vγ9Vδ2 T cell activation. link. Fortunately, new CryoEM studies from two Chinese groups, support this model link link
We are currently continuing the analyses of the mechanism of activation of Vγ9Vδ2 T cells, which serve the development of a Vγ9Vδ2 T cell-based tumor therapy. A new field of research is the modulation of the immune response by BTN(3) molecules. Here we can draw on our experience in the analysis of the molecular basis of BTN-dependent Vγ9Vδ2 T cell activation. link